

成都生物所研究揭示珍稀瀕危物種鱷蜥的體色多態(tài)性及其信息傳遞功能
作者:邱霞
時(shí)間:2022-09-26
動(dòng)物體色多態(tài)性及其進(jìn)化是當(dāng)前進(jìn)化生物學(xué)領(lǐng)域研究的熱點(diǎn)問(wèn)題之一(Endler et al., 2017;Stuart-Fox et al. 2020)。動(dòng)物體色為何如此多變?導(dǎo)致體色多變的選擇壓力是什么?體色變化和個(gè)體身體狀態(tài)、行為策略、社會(huì)地位等的相互關(guān)系是什么?體色多態(tài)性的維持機(jī)制是什么?這一系列問(wèn)題正在讓動(dòng)物體色成為進(jìn)化生物學(xué)多分支學(xué)科共同關(guān)注的焦點(diǎn)。
體色變異是體色多態(tài)性形成的前提(Tibbetts et al. 2017)。經(jīng)典理論認(rèn)為,動(dòng)物體色變異主要源于自然選擇和性選擇,前者主要強(qiáng)調(diào)體色變異與個(gè)體生存的相互關(guān)系,即動(dòng)物通過(guò)體色變異,可以提升自己的偽裝效果,從而降低被天敵捕食的風(fēng)險(xiǎn);后者主要強(qiáng)調(diào)體色變異與競(jìng)爭(zhēng)配偶的相互關(guān)系,即動(dòng)物通過(guò)體色變異能向競(jìng)爭(zhēng)對(duì)手傳遞自己的身體狀況和社會(huì)地位信息,從而在同性競(jìng)爭(zhēng)和異性選擇過(guò)程中占據(jù)優(yōu)勢(shì)。現(xiàn)代進(jìn)化生物學(xué)理論認(rèn)為:動(dòng)物體色變異的形成可能是社會(huì)選擇的結(jié)果,主要影響社會(huì)交流的過(guò)程,因?yàn)楹芏鄤?dòng)物體色變異的形成與生存和競(jìng)爭(zhēng)配偶似乎沒(méi)有直接的聯(lián)系,比如雌性個(gè)體在繁殖后期的體色變異。無(wú)論是經(jīng)典理論,還是現(xiàn)代理論,絕大多數(shù)關(guān)于體色變異的研究都是基于雄性個(gè)體,關(guān)于雌性個(gè)體體色變異的研究才剛剛起步。
鱷蜥(Shinisaurus crocodilurus)是研究體色變異的良好模型,與其它蜥蜴相比,雄性和雌性鱷蜥喉部和腹部都表現(xiàn)出不同的顏色(圖1)。何南等(2011)首次報(bào)道了鱷蜥的性二色型,成年雄性鱷蜥頭胸腹面是淺藍(lán)色或鮮紅色,而雌性是淺黃色或淡紅色。Van Schingen等(2016)也報(bào)道了鱷蜥喉部顏色的多態(tài)性,包括藍(lán)色、紅色和黃色。實(shí)際上,鱷蜥的體色變異可能比已有報(bào)道更豐富,喉部和腹部淺色或深色背景上有許多鮮艷的橙色或紅色色斑。對(duì)蜥蜴類動(dòng)物而言,喉部與腹側(cè)色斑通常被用作社會(huì)交流的信號(hào),能夠反映個(gè)體的身體質(zhì)量狀態(tài)或行為策略信息(Tibbetts et al. 2017)。因此,研究雌性和雄性鱷蜥色斑變異在社會(huì)交流中的功能對(duì)理解體色多態(tài)性的進(jìn)化,特別是雌性體色多態(tài)性的進(jìn)化具有重要意義。此外,探明鱷蜥色斑多態(tài)性及其信息傳遞功能,也有助于指導(dǎo)該物種的圈養(yǎng)繁殖和野外放歸。由于國(guó)際寵物貿(mào)易和棲息地退化的威脅,該物種已被列為國(guó)家一級(jí)保護(hù)動(dòng)物,CITES附錄I物種,并于2014年被列入IUCN紅色名錄瀕危物種。解析該物種色斑多態(tài)性及其與個(gè)體質(zhì) 量的相關(guān)關(guān)系,有助于建立一種高效快捷的個(gè)體評(píng)估方法,并促進(jìn)人工繁育和野外放歸群體設(shè)計(jì)。
量的相關(guān)關(guān)系,有助于建立一種高效快捷的個(gè)體評(píng)估方法,并促進(jìn)人工繁育和野外放歸群體設(shè)計(jì)。
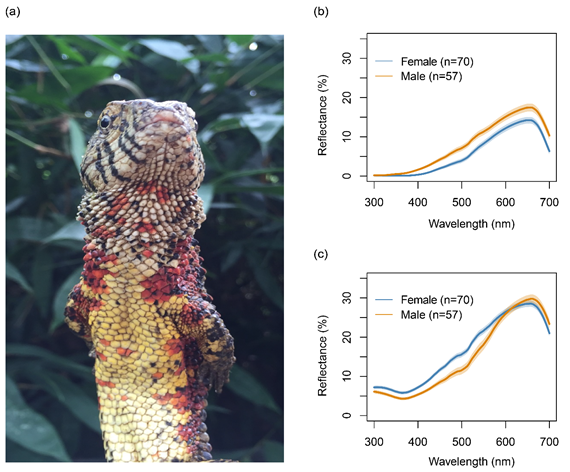
圖1(a)鱷蜥喉部和腹側(cè)色斑展示;(b)雌性和雄性鱷蜥喉部色斑的光譜反射曲線;(c)雌性和雄性腹側(cè)色斑的光譜反射曲線。
我們采用便攜式光譜儀量化了鱷蜥喉部和腹部的體色變化,研究目的包括:(1)量化該物種的體色變異,判定不同部位的色斑是否能被劃分為不同的色型;(2)采用視覺(jué)建模方法計(jì)算不同色斑在蜥蜴視覺(jué)系統(tǒng)感受下的明顯程度,判斷其能被同類識(shí)別的程度;(3)量化體色變異與個(gè)體質(zhì)量(身體狀況、咬合力等)和繁殖力(產(chǎn)仔數(shù)和窩仔質(zhì)量等)之間的關(guān)系。本研究共涉及127只成年鱷蜥,所用個(gè)體均來(lái)自廣西大桂山鱷蜥國(guó)家級(jí)自然保護(hù)區(qū)(北緯24°09′,東經(jīng)111°81′)鱷蜥繁殖中心。
研究結(jié)果顯示:雌性和雄性鱷蜥喉部和腹側(cè)色斑都可以劃為兩種類型,即紅色色斑和非紅色斑(圖2)。喉部色斑與腹部色斑無(wú)必然聯(lián)系,喉部為紅色的個(gè)體,腹部可能為非紅色,此外色斑類型與性別無(wú)關(guān),雄性和雌性個(gè)體可能擁有相似的色斑變異。有趣的是,相同個(gè)體的色斑類型隨著年齡而發(fā)生變化,出生時(shí)為白色的個(gè)體,在成體階段可能保持白色,或者變?yōu)榧t色,出生時(shí)為紅色的個(gè)體,其色斑可能終身維持。
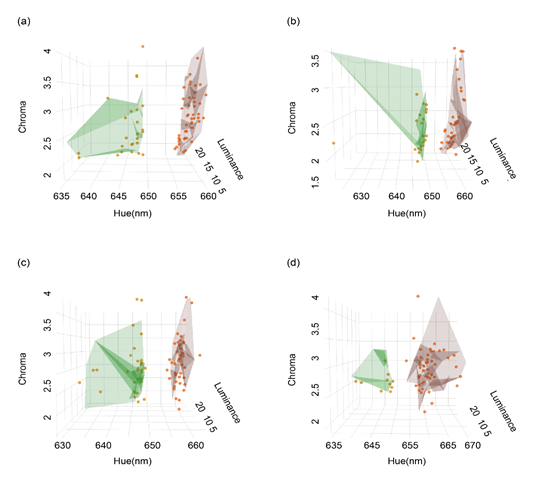
圖2通過(guò)光譜參數(shù)對(duì)鱷蜥不同性別的喉部和腹側(cè)色斑的聚類分析。(a)雌性喉部色斑類型;(b)雄性喉部色斑類型;(c)雌性腹側(cè)色斑類型;(d)雄性腹側(cè)色斑類型。
其次,參考蜥蜴視覺(jué)系統(tǒng)進(jìn)行視覺(jué)建模,結(jié)果顯示:喉部和腹部色斑的可區(qū)分度(JND)均大于蜥蜴可區(qū)分閾值1,說(shuō)明兩種類型的色斑均能被同類識(shí)別(圖3a)。其次,蜥蜴對(duì)喉部?jī)煞N色斑之間的區(qū)分度也大于其可區(qū)分閾值1JND(圖3b和c),表明蜥蜴能識(shí)別不同的色斑類型。綜上,鱷蜥的色斑在社會(huì)交流過(guò)程中發(fā)揮重要作用。
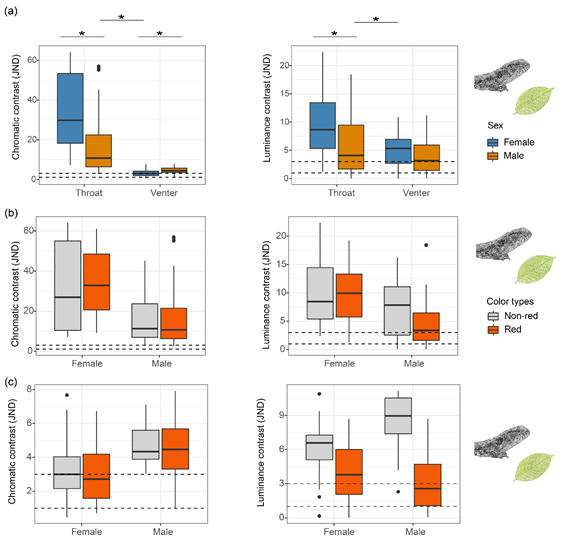
圖3(a)鱷蜥喉部和腹側(cè)色斑在蜥蜴視覺(jué)系統(tǒng)下的區(qū)分度(JND);(b)喉部和(c)腹部紅色色斑與非紅色斑類型之間的可區(qū)分度。“*”表示線性混合模型檢測(cè)到顯著性差異。
色斑類型與個(gè)體質(zhì)量相關(guān)性分析顯示:鱷蜥色斑能夠傳遞個(gè)體質(zhì)量信息(圖4)。喉部呈紅色的雌性咬合力大,而喉部呈紅色的雄性年齡大。咬合力與年齡通常是個(gè)體身體質(zhì)量的指標(biāo),咬合力大的蜥蜴在競(jìng)爭(zhēng)配偶時(shí)獲勝幾率更高(Healey et al., 2007);年齡大的動(dòng)物往往有更多的求偶和覓食經(jīng)驗(yàn),有利于提高生存和繁殖適合度。此外,腹部呈紅色的雌性頭部更大。頭部大小往往代表咬肌的發(fā)達(dá)程度(Huyghe et al., 2008),使紅色的蜥蜴在競(jìng)爭(zhēng)配偶時(shí)獲勝幾率更高(Healey et al., 2007)。我們還發(fā)現(xiàn)了其他與色斑類型有關(guān)的形態(tài)學(xué)差異。喉部呈紅色的雌性身體細(xì)長(zhǎng),且尾巴較長(zhǎng),而喉部無(wú)紅色的雌性則較粗壯,尾巴較短,表明鱷蜥的色斑類型可能代表了某種行為對(duì)策,細(xì)長(zhǎng)的體型對(duì)樹棲的鱷蜥和通過(guò)游泳逃生的鱷蜥有著重要的意義。
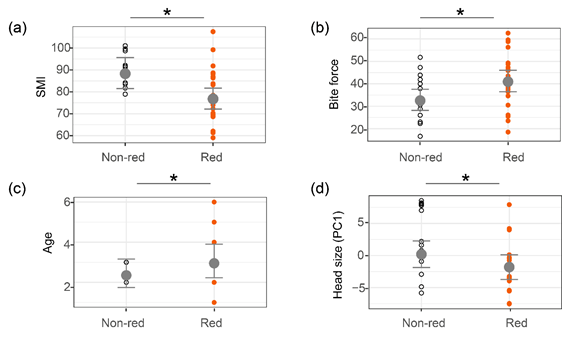
圖4喉部和腹側(cè)色斑類型與個(gè)體質(zhì)量之間的相關(guān)關(guān)系。“*”表示線性混合模型檢測(cè)到的顯著性差異。(a)SMI和(b)咬合力在雌性喉部色斑類型間的差異;(c)雄性年齡在喉部色斑類型間差異;(d)雌性頭大小在腹側(cè)色斑類型間的差異。
色斑類型與個(gè)體繁殖能力相關(guān)性分析顯示:喉部和腹部色斑與雌性的繁殖力無(wú)關(guān)。但是腹側(cè)色斑明顯的雄性個(gè)體后代更大,且存活率更高(圖5)。這提示,在未來(lái)規(guī)劃鱷蜥的人工繁育和放歸計(jì)劃時(shí),應(yīng)該充分考慮不同體色個(gè)體的組成,腹部紅色的雄性個(gè)體可能作為放歸個(gè)體的主要成分。
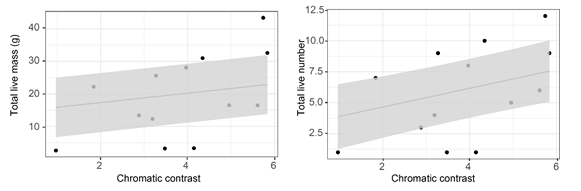
圖5雄性腹側(cè)色斑顏色明顯程度與存活子代質(zhì)量和數(shù)量的相關(guān)關(guān)系。
研究結(jié)果以“Colour Variation in the Crocodile Lizard (Shinisaurus crocodilurus) and Its Relationship to Individual Quality”為題發(fā)表于國(guó)際科學(xué)期刊《Biology》。中國(guó)科學(xué)院成都生物研究所動(dòng)物行為與仿生項(xiàng)目組邱霞博士為該論文第一作者,齊銀副研究員為通訊作者;澳大利亞麥考瑞大學(xué)Martin J. Whiting教授、加拿大圭爾夫大學(xué)傅金鐘教授、中國(guó)科學(xué)院動(dòng)物研究所杜衛(wèi)國(guó)研究員、廣西師范大學(xué)武正軍教授和廣西大桂山鱷蜥自然保護(hù)區(qū)羅樹毅參與了該項(xiàng)目。本研究得到國(guó)家自然科學(xué)基金(31872233,31821001)、四川省杰出青年項(xiàng)目(2020JDJQ0001)和西藏自治區(qū)科學(xué)與技術(shù)計(jì)劃(XZ202201ZY0030G)的資助。
參考文獻(xiàn):
何南, 武正軍, 蔡鳳金, 等, 2011. 鱷蜥的兩性異形[J]. 生態(tài)學(xué)雜志, 30(01): 7.
ENDLER J A, MAPPES J, 2017. The current and future state of animal coloration research[J/OL]. Philosophical Transactions of the Royal Society of London B Biological Sciences, 372(1724). DOI:10.1098/rstb.2016.0352.
HEALEY M, ULLER T, OLSSON M, 2007. Seeing red: morph-specific contest success and survival rates in a colour-polymorphic agamid lizard[J/OL]. Animal Behaviour, 74(2): 337-341. DOI:10.1016/j.anbehav.2006.09.017.
HUYGHE K, HERREL A, ADRIAENS D, et al., 2008. It is all in the head: morphological basis for differences in bite force among colour morphs of the Dalmatian wall lizard[J/OL]. Biological Journal of the Linnean Society, 96(1): 13-22. DOI:10.1111/j.1095-8312.2008.01103.x.
STUART-FOX D, AULSEBROOK A, RANKIN K J, et al., 2020. Convergence and divergence in lizard colour polymorphisms[J]. Biological Reviews: 21.
TIBBETTS E A, MULLEN S P, DALE J, 2017. Signal function drives phenotypic and genetic diversity: the effects of signalling individual identity, quality or behavioural strategy[J/OL]. Philosophical Transactions of the Royal Society B: Biological Sciences, 372(1724): 20160347. DOI:10.1098/rstb.2016.0347.
